Howell G.M. Edwards,*a,b Ian B. Hutchinson,b Richard Ingley,b Nick R. Waltham,c Sarah Beardsley,c Shaun Dowsonc and Simon Woodwardc
aCentre for Astrobiology & Extremophiles Research, School of Life Sciences, University of Bradford, Bradford, West Yorkshire BD7 1DP, UK. E-mail: [email protected]
bDepartment of Physics and Astronomy, Space Research Centre, University of Leicester, University Road, Leicester LE1 7RH, UK
cImaging Systems Division, Space Science and Technology Department, Science and Technology Facilities Council, Rutherford Appleton Laboratory, Harwell Science and Innovation Campus, Didcot, Oxfordshire OX11 0QX, UK
Introduction
The discovery of water and ice on Mars by NASA (National Aeronautics and Space Administration) rover vehicles and orbiters and the appreciation that early Mars probably developed in a similar way to early Earth has focused attention on the biochemical survival strategies adopted by terrestrial extremophiles, which exist in a range of geological niches and habitats having a parallel on Mars, termed terrestrial Mars analogue sites. The European Space Agency Aurora programme, and in particular the ExoMars/Pasteur mission which will search for extinct or extant life signatures on Mars, has identified several analytical instruments to assess the biological status of the planet before human exploration of the Martian surface will be undertaken in the next few decades. Raman spectroscopy has been selected as one of the first-pass analytical instruments for this ExoMars mission to identify the geological and biogeological spectral signatures that could herald the presence of extinct or extant life on Mars. This article describes the type of information that Raman spectroscopic studies of terrestrial extremophiles has provided about their protective suites of biochemicals synthesised to combat their exposure to harsh environments at “limits of life” situations; the recognition of key biomolecular spectral markers in the geological record provides the basis of a database of life detection both terrestrially and extraterrestrially. The advantages of Raman spectroscopy for the detection of biomolecules in geological mineral matrices will be illustrated using terrestrial extremophilic specimens from hot and cold deserts where temperatures, desiccation and exposure to low wavelength, high energy radiation in the ultraviolet region demand response strategies for species survival.
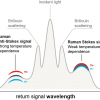
The application of Raman spectroscopic techniques to the characterisation of the protective biochemicals used in the survival strategies of extremophilic organisms in terrestrially stressed environments,1,2 coupled with the palaeogeological recognition that early Mars and Earth had maintained similar environments under which Archaean cyanobacteria could have developed,3 has driven the proposal for the adoption of Raman spectroscopy as novel analytical instrumentation for planetary exploration.4,5,6 The announcement by the European Space Agency (ESA) that a miniaturised Raman spectrometer would form part of the Pasteur analytical life-detection protocol in the ExoMars mission for the search for traces of life on Mars in the Aurora space exploration programme has confirmed that Raman spectroscopy will perform a key role for the molecular and elemental analysis of the Martian surface and subsurface on the ExoMars rover vehicle (Figure 1, Table 1).

Table 1. Instruments on the ESA ExoMars Rover, with national PI interest in brackets.
| PanCam: Panoramic Camera—digital terrain mapping (UK, D, Sui) MOMA: Mars Organic Molecule Analyser—GC/MS system for detection of volatiles in atmosphere and sediments (D, F, USA) MicroOmega-IR: Infrared imaging spectrometer (F, Sui) MARS-XRD: X-ray diffractometer for minerals (I, UK) RLS: Raman laser spectrometer—geological and biogeological molecules (E, F) WISDOM: ground penetrating radar for water ice, subsurface deposits (F, N, D) MA-MISS: Mars multi-spectral imager CLUPI: close-up imager—high-resolution camera for rocks, drill cores and powders (Sui, F) LMC: life marker chip—detection of specific molecules associated with life (UK) |
It is undeniable that the most important scientific discovery in a future space mission would be evidence for life on another planet, whether this be extant or extinct; however, this statement itself generates two philosophical questions, namely, how do we define life and how would we then recognise it or its residues in the planetary geological record? We must also address the possibility that any extraterrestrial organism identified in space exploration could have originated on Earth and have been transported to our planetary neighbours either by our own intervention, reinforcing the need for planetary protection protocols for spacecraft and landers, or through panspermia. The precise definition of life is rather elusive and many attempts to do so have been eventually deemed unsatisfactory;7,8,9,10 the NASA definition of life as A self-sustaining system capable of Darwinian evolution” incorporates a molecular genesis with replicative procedures and avoids several pitfalls of alternative definitions based upon an ability of the system to reproduce.11
At this juncture, therefore, it is appropriate to consider some of the scientific parameters that will need to be considered for the detection of life in the ExoMars project; primarily, the selection criteria for an analytical astrobiological mission such as ExoMars need to consider the following questions:
- What organisms could have existed and possibly survived the current and past extremes of environment on Mars?
- What type of geological niches is to be found which may conceal the traces of relict or extant life on Mars?
- What signatures would these organisms have left in such environments as indicators of their presence and how are we going to recognise them?
- What molecules could be considered as constituting a proof of life on Mars?
- Are there terrestrial scenarios or putative Mars analogue sites which could be used as “models” for the evaluation of these scientific questions?
The historical Mars
From the birth of our Solar System some 4.6 Gya, the terrestrial geological record suggests that microbial autotrophic ecosystems already existed on Earth from 3.5 Gya to 3.8 Gya. There is now much evidence that early Earth and early Mars were indeed very similar in their physico–chemical composition; since Mars is significantly smaller than Earth, it is very likely that planetary cooling occurred more rapidly and that Mars was able to sustain an aqueous environment on its surface earlier than was possible on Earth. In the Epoch I (ca 4.65–3 Gya) period of Mars proposed by McKay,3 the planet was probably more temperate and wet and since there is geological evidence that life had already started on Earth during this period, it seems reasonable to conclude that life could also have started on Mars too; by Epoch IV (ca 1.5 Gya to present), however, catastrophic changes on Mars would have compromised the survival of organisms on the Martian surface and it is possible that the Martian analogues of terrestrial extremophiles would have been the last survivors of life on Mars through their environmental adaptation in Martian geological niche sites.
Analytical astrobiology of Mars
The detection of biomolecular markers in geological substrates or the subsurface regolith of Mars is a primary goal for astrobiology;12,13 however, the evolutionary pressure of environmental stress on Mars, especially the high levels of low wavelength ultraviolet radiation insolation, low temperatures, extreme desiccation and hypersalinity would have demanded appropriately severe protective strategies to promote the origin, survival and evolution of microbial life.14 The ultraviolet radiation protection afforded to subsurface organisms by the iron (III) oxide surface regolith acting as a filter has been proposed as a key factor for the maintenance of biomolecular activity at the Martian surface,15 but the same ultraviolet and low wavelength electromagnetic radiation insolation generates hydroxyl radicals and peroxides in the surface regolith which would certainly inhibit the survival of complex biomolecules in the surface oxidation zone. In this respect the diagnosis, catagenesis and biodegradation of terrestrial Mars analogues to give the fossils recognisable in our own geological record would not be transposable to a Martian scenario. Hence, the complex chemical systems comprising terrestrial soils, bitumens and kerogens found in our own planetary lithology16,17,18,19 and which provide much valuable information about their sourcing processes would not be expected to occur to the same extent on Mars.
However, it is believed that Mars might still preserve a chemical record of early life in rocks from the Noachian era, which overlaps the terrestrial Archaean geological history, from about 3.8 Gya. The search for extinct or extant life on Mars must therefore centre upon the identification and recognition of specially protected niche geological sites in which the biomolecular signatures would be preserved; the fundamental approach must then consider the detection of key molecular biomarkers, probably within rocks and certainly subsurface, perhaps even in ancient lacustrine sediments,20,21 which will necessitate the deployment of remote analytical sensors with preset protocols and an established database recognition strategy for minerals, biologically modified geological strata and biomolecular residues. Examples from terrestrial analogue sites include carbonates, carbonated hydroxyfluoroapatite, gypsum, calcium oxalates, porphyrins, carotenoids, scytonemin and anthraquinones.22,23
Clearly, the identification and selection of terrestrial Mars analogue sites19,20 will be a critical step in the development of an analytical astrobiology for Mars in two respects; first, the understanding of the type of geological formations that have been colonised by extremophilic organisms in terrestrial “limits of life” situations and habitats,20 and second, the deployment of novel analytical instrumentation which can reveal the presence of the key signatures of extinct and extant life in micro-niches in the geological record.21,22,23,24,25 The gathering of data from such terrestrial Mars analogue sites is therefore the first step to be taken in the search for extraterrestrial life signatures; in this, the application of Raman spectroscopic techniques has been successful through the direct characterisation of the molecular and molecular ionic signatures of biomolecules and their modified structures situated in the geological record which does not involve either the physical or chemical separation of the organic and inorganic components. Some of these terrestrial Mars analogue sites are described below and the data obtained using laboratory-based Raman spectroscopic techniques have advanced our understanding of extremophile behaviour significantly.
In a recent special issue of the Philosophical Transactions of the Royal Society, in a year that celebrated its 350th anniversary as the longest running scientific journal, several articles26,27,28,29,30,31,32,33 highlighted the role of Raman spectroscopy in the characterisation of biosignatures of extremophilic colonisation of geological substrates in a range of stressed terrestrial environments; these articles and others in the same issue which address the detection of geological and biogeological spectral markers that are relevant to space missions34,35,36,37 give a very good appreciation of the spectroscopic requirements that will be essential for the construction of a relevant spectral database for the ExoMars and forthcoming space missions which have a Raman spectrometer aboard their rover vehicles. Some selected examples of the data which can be provided by the Raman spectroscopic interrogation of terrestrial Mars analogue sites will now be highlighted.
Terrestrial Mars analogue sites
Andrews Ridge, Long-Term Ecological Research (LTER) Site, Lake Hoare, Taylor Valley, Antarctica
A specimen of igneous feldspar shows several stratified deposits; in particular, two green-coloured zones situated some 10–15 mm beneath the surface provide an example of chasmolithic extremophilic colonisation. The substrate has a Raman spectrum characteristic of the minerals quartz (bands at 465, 356, 206 and 127 cm–1) and microclinic feldspar (with characteristic bands at 512 cm–1 and 155 cm–1). The Raman spectrum of the chasmolithic zone shows bands due to aragonite (1086, 705, 207 and 154 cm–1) and can be seen along with those of quartz and microclinic feldspar (plagioclase). The presence of a beta-carotene biomarker is evidenced by the bands at 1518, 1156 and 1003 cm–1 and confirms the biological component in this zone. The detection of aragonite in this zone and not elsewhere in the substratum is also indicative of past biological activity, making it a biogeomarker in this context, which can be correlated with the geological and sedimentary history of the LTER site.38
Mars Oasis and Battleship Promontory, Alatna Valley, Antarctica
As the environmental habitat progressively worsens towards higher Polar latitudes, the survival strategies of biological organisms require further adaptation until at the “limits of life scenario” it is impossible for life to survive in an epilithic form and then the surviving biology becomes endolithic. The requirement for photosynthetically active radiation with the necessity for effective protection from the low wavelength UV radiation demands the synthesis of a sophisticated suite of biochemicals and accessories which can assist the host organism in its survival strategy and the adaptation of the specific geology to assist this necessity. A typical endolith in Beacon sandstone is shown in Figure 2(a) from Mars Oasis, Antarctica, where the stratified components comprising the crustal rock and the iron (III) oxide depleted layer containing the dark greenish-black Chroococcidiopsiscyanobacterial colonies can be seen. Raman spectroscopy has been successful in the provision of information about these systems since a key part of the survival strategy depends on the molecular interactions between the host mineral substrate and the cyanobacteria and data can be obtained non-destructively. Other analytical methods hitherto have necessitated the physical and chemical separation of the inorganic and biological components, whereby much interactive information is lost. The Raman spectrum shown in Figure 3 indicates the presence of chlorophyll, calcium oxalate monohydrate and a carotenoid, which are all essential chemicals produced for the photosynthesis, radiation photoprotection and anti-desiccation response of the organism in its extremophilic environment along with the quartz and haematite components of the geological matrix. A further biomodification of the geological substrate which is essential for the survival of the colony is the transportation of iron (III) oxide to the rock surface, and this can be in the depletion of pigment from the colonisation zone seen in Figure 2(b); the Raman spectrum of the surface of the Battleship Promontory specimen, provides the supporting information that a change has been effected in the composition of the iron (III) oxide during this process and signatures of goethite, an iron (III) oxide hydroxide, can now be noted at the upper surface of the sample containing the endolith, which is absent from specimens of rock that are not colonised. Clark15 has demonstrated that the increased protection afforded to endolithic colonies effected through this transportation of the iron (III) oxide can materially assist their ability to filter out UVB radiation from the insolation at the rock surface. The significant presence of iron (III) oxide at the surface of Mars is therefore of considerable interest and relevance to the possibility of protection being afforded to the survival of organisms under the surface of Mars, which will be addressed in the ExoMars mission through the provision of a drill for sampling the subsurface regolith.
 Endolithic specimen of Chroococcidiopsis cyanobacteria in Beacon Sandstone from Mars Oasis, Antarctica; (b) Beacon Sandstone from Battleship Promontory, Alatna Valley, Antarctica, demonstrating the depletion zone of iron (III) oxide and the colour change at the surface resulting from the conversion of haematite to goethite. Reproduced courtesy ESA.")

Haughton Crater, Devon Island, Canadian High Arctic
This is a meteorite crater dating from 26 Mya, where the impact created selenite (a transparent form of gypsum, CaSO4.2H2O) crystals within the melt breccia deposits at the rim of the crater; several crystals are found to contain dark brown–black inclusions at depths of up to 5 cm below the surface, which prove to be examples of colonies of halotrophic (salt-tolerating) extremophiles. One of these inclusions, along with its accompanying Raman spectrum (Figure 4) which was recorded confocally and directly through the transparent crystal without effecting chemical or mechanical pretreatment, identifies the colony as Nostoc cyanobacteria39 through its characteristic suite of protective biochemicals; the confocal microRaman spectra of other inclusions has identified colonies of a Nostoc species.

Volcanic Basalt, Scott-Keltie Fiellet Ridge, Spitzbergen, Arctic Norway
In this case study several different types of extremophile can be identified in the same specimen where an endolithic colony is found inside a vacuole in the basalt connected to the surface via a small orifice.40 Fracturing the vacuole exposes the endolithic colony inside the cavity (Figure 5). The Raman spectrum of this colony with 785 nm excitation clearly shows the signatures of lutein, a carotenoid, chlorophyll and c-phycocyanin, a light-harvesting accessory pigment. With 514.5 nm excitation, however, the increased selectivity for detection of carotenoids through the operation of a resonance Raman effect at this wavelength demonstrates the presence of two different carotenoids, namely astaxanthin and lutein, in the endolithic colony (Figure 6).


Nakhla SNC Martian Meteorite
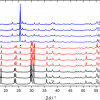
A fragment of the Nakhla SNC (Shergotty-Nakhla-Chassigny) Mars meteorite, one of the 30 or so Martian meteorites that have now been identified, that fell in Egypt in 1911 has been analysed using Raman spectroscopy and a series of stack-plotted spectra are shown in Figure 7. The spectra show a diversity of composition and heterogeneity in this specimen, where the sampling regions range, from a to d respectively, from clear glassy, creamy-white crystalline, to white crystalline and black glassy areas in the specimen. Bands due to feldspar, silicates and olivine are identifiable; a noteworthy feature in spectrum 8b is a very weak band arising from calcium carbonate at 1086 cm–1 and this provides a focus of attention for the possible presence of carbonates as geological deposits on Mars.41

Conclusions
Lichens and cyanobacteria are pioneer colonisers of extreme terrestrial environments and their strategic adaptation for survival in lithic substrata through their ability to adapt to and to modify their geological habitats provides a new insight into extremophilic behaviour. A recent focus for astrobiology is the remote Raman spectroscopic sensing of planetary surfaces for the presence of key biomolecular markers in the search for extant or extinct life, the last surviving vestiges of which could well be analogous to the organisms and scenarios that have been reported here and comprising endolithic and chasmolithic colonies at the “limits of life” terrestrially; a major advantage of Raman spectroscopy in this respect is the ability to record data from geological and biological components of a specimen which is essential for extremophilie characterisation.
The Arctic and Antarctic cold deserts, in particular, provide examples of some of the most extreme terrestrial habitats for extremophilic organisms, where there are large seasonal fluctuations in surface temperatures, high levels of desiccation, strongly abrasive katabatic winds and intense solar radiation insolation which is increased in low wavelength, high energy UVB and UVC components during the spring depletion of protective atmospheric ozone at high latitudes. Survival strategies for cyanobacteria under these conditions involves the biogeological modification of their habitats as well as the synthesis of specialised suites of protective biochemicals, many of which apparently have a duality of function in protection of the colonies and damage repair mechanisms. The identification of the key Raman spectroscopic biosignatures belonging to these biomolecular protectants and auxiliary species is an essential criterion for the construction of a Raman spectral database that is proposed for the indication of extant or extinct life in rocks. The compilation of such a database is a critical stage in the provision of miniaturised life-detection instrumentation based on Raman spectroscopy and destined for incorporation into robotic planetary landers and for the evaluation of data retrieved from these missions; this article has described some terrestrial scenarios that are relevant to extraterrestrial exploration for life signatures and will form part of an expanding database for the recognition of early evolutionary processes on planetary surfaces and subsurfaces.
The advantages of Raman spectroscopy for the identification of life signatures in extreme environments can thus be summarised as follows:
- Specific biomolecular fingerprinting based upon key indicators such as carotenoids, chlorophyll, aromatic hydrocarbons, porphyrins and oxalates.
- Remote analytical ability using fibre optical probes.
- Surface preparation not essential.
- Spatial resolution to the micron level.
- Spectral range covers most signatures from inorganic substrates and their biologically modified forms and organic protectant biomolecules in geological matrices.
- No interference from water and ice, which is relevant for permafrost and polar regions where life signatures may be sought beneath glacial sites.
Key Raman spectral features for geological and biological markers that could form a preliminary database for the recognition of minerals and biomarkers on Mars is given in Table 2; the diverse range of relevant materials that have definitive Raman bands should be noted.
Calcite | CaCO3 | 1086 | 712 | 282 | 156 |
|
|
|
|
|
|
|
|
|
|
Aragonite | CaCO3 | 1086 | 704 | 208 | 154 |
|
|
|
|
|
|
|
|
|
|
Dolomite | CaMg(CO3)2 | 1098 | 725 | 300 | 177 |
|
|
|
|
|
|
|
|
|
|
Magnesite | MgCO3 | 1094 | 738 | 330 | 213 | 119 |
|
|
|
|
|
|
|
|
|
Hydromagnesite | Mg5(CO3)4(OH)2.4H2O | 1119 | 728 | 326 | 232 | 202 | 184 | 147 |
|
|
|
|
|
|
|
Gypsum | CaSO4.2H2O | 1133 | 1007 |
| 669 | 618 | 492 | 413 |
|
|
|
|
|
|
|
Anhydrite | CaSO4 | 1015 | 674 | 628 | 500 | 416 |
|
|
|
|
|
|
|
|
|
Quartz | SiO2 | 1081 | 1064 | 808 | 796 | 696 | 500 | 542 | 463 | 354 | 263 | 206 | 128 |
|
|
Haematite | Fe2O3 | 610 | 500 | 411 | 293 | 245 | 226 |
|
|
|
|
|
|
|
|
Limonite | FeO(OH)nH2O | 693 | 555 | 481 | 393 | 299 | 203 |
|
|
|
|
|
|
|
|
Apatite | Ca5(PO4)3(F,Cl,OH) | 1034 | 963 | 586 | 428 |
|
|
|
|
|
|
|
|
|
|
Weddellite | Ca(C2O4)2H2O | 1630 | 1475 | 1411 | 910 | 869 | 597 | 506 | 188 |
|
|
|
|
|
|
Whewellite | Ca(C2O4)H2O | 1629 | 1490 | 1463 | 1396 | 942 | 896 | 865 | 596 | 521 | 504 | 223 | 207 | 185 | 141 |
Chlorophyll | C55H72O5N4Mg | 1438 | 1387 | 1326 | 1287 | 1067 | 1048 | 988 | 916 | 744 | 517 | 351 |
|
|
|
c-phycocyanin | C36H38O6N4 | 1655 | 1638 | 1582 | 1463 | 1369 | 1338 | 1272 | 1241 | 1109 | 1054 | 815 | 665 | 499 |
|
Beta-carotene | C40H56 | 1515 | 1155 | 1006 |
|
|
|
|
|
|
|
|
|
|
|
Rhizocarpic acid | C26H23O6 | 1665 | 1610 | 1595 | 1518 | 1496 | 1477 | 1347 | 1303 | 1002 | 944 | 902 | 787 | 448 |
|
Scytonemin | C36H20N2O4 | 1605 | 1590 | 1549 | 1444 | 1323 | 1283 | 1245 | 1172 | 1163 | 984 | 752 | 675 | 574 | 270 |
Calycin | C18H10O5 | 1653 | 1635 | 1611 | 1595 | 1380 | 1344 | 1240 | 1155 | 1034 | 960 | 878 | 498 | 484 |
|
Parietin | C16H12O5 | 1671 | 1631 | 1613 | 1153 | 1387 | 1370 | 1277 | 1255 | 926 | 571 | 519 | 467 | 458 | 398 |
Usnic acid | C18H16O7 | 1694 | 1627 | 1607 | 1322 | 1289 | 1192 | 1119 | 992 | 959 | 846 | 602 | 540 |
|
|
Emodin | C15H10O5 | 1659 | 1607 | 1577 | 1557 | 1298 | 1281 | 942 | 565 | 467 |
|
|
|
|
|
Atranorin | C19H18O8 | 1666 | 1658 | 1632 | 1303 | 1294 | 1266 | 588 |
|
|
|
|
|
|
|
Pulvinic dilactone | C18H10O4 | 1672 | 1603 | 1455 | 1405 | 1311 | 981 | 504 |
|
|
|
|
|
|
|
Gyrophoric acid | C24H20O10 | 1662 | 1628 | 1612 | 1334 | 1304 | 1291 | 1235 | 1138 | 561 |
|
|
|
|
|
Acknowledgements
The authors express their gratitude to colleagues in the European Space Agency ExoMars Raman RLS Science and Instrument Teams for interdisciplinary expertise in the achievement of acceptance of miniaturised instrumentation for the life-detection mission to Mars and to the UK Science and Technology Facilities Research Council for the ongoing funding support for Raman studies relevant to early evolution and the ExoMars mission.
References
- D.D. Wynn-Williams and H.G.M. Edwards, Planet. Space Sci. 48, 1065–1075 (2000).
- D.D. Wynn-Williams and H.G.M. Edwards, Icarus 144, 486–503 (2000).
- C.P. McKay, Origins Life Evol. B. 27, 263–289 (1997).
- H.G.M. Edwards and E.M. Newton, “Applications of Raman spectroscopy for exobiological prospecting”, in The Search for Life on Mars, Ed by J.A. Hiscox. British Interplanetary Society, London, pp. 75–83 (1999).
- H.G.M. Edwards, “Analytical Astrobiology: The Role of Raman Spectroscopy in Biomarker Speciation for Extraterrestrial Life Detection”, in Astrobiology: Emergence, Search and Detection of Life, Ed by V.A. Basiuk. American Scientific Publishers, Stevenson Ranch, CA, USA, pp. 361–375 (2010).
- H.G.M. Edwards, E.M. Newton, D.D. Wynn-Williams, D.L. Dickensheets, C. Schoen and C. Crowder, Intl. J. Astrobiol. 1, 333–348 (2003).
- C.E. Cleland and C.F. Chyba, Origins Life Evol. B. 32, 387–393 (2002).
- D.E. Koshland, Science 295, 2215–2216 (2002).
- S.A. Benner, Astrobiology 10, 1021–1030 (2010).
- S. Tirard, M. Morange and A. Lazcano, Astrobiology 10, 1003–1009 (2010).
- G.F. Joyce, Foreword, in Origins of Life: the Central Concepts, Ed by D.W. Deamer and G. Fleischaker. Jones and Bartlett, Boston, USA (1994).
- H.G.M. Edwards, Origins Life Evol. B. 34, 3–11 (2004).
- H.G.M. Edwards, C.D. Moody, S.E. Jorge Villar and D.D. Wynn-Williams, Icarus 174, 560–571 (2005).
- C.S. Cockell and J.R. Knowland, Biol. Revs. 74, 311–345 (1999).
- B.C. Clark, J. Geophys Res.-Planet. 103, 28545–28556 (1998).
- J. Jehlicka, H.G.M. Edwards and P. Vitek, Planet. Space Sci. 57, 606–613 (2009).
- D. Pullan, F. Westall, B.A. Hofmann, J. Parnell, C.S. Cockell, H.G.M. Edwards, S.E. Jorge Villar, C. Schröder, G. Cressey, L. Marinangeli, L. Richter and G. Klingelhöfer, Astrobiology 8(1), 119–156 (2008).
- H.G.M. Edwards, S.E. Jorge Villar, D. Pullan, B.A. Hofmann, M.D. Hargreaves and F. Westall, J. Raman Spectrosc. 38, 1352–1361 (2007).
- S.E. Jorge Villar, L.G. Benning and H.G.M. Edwards, Geochem. Trans. 8(8), 1–11 (2007).
- J.L. Bishop, B.L. Aglen, L.M. Pratt, H.G.M. Edwards, D.J. des Marais and P.T. Doran, Intl. J. Astrobiol. 2, 1–15 (2003).
- P.T. Doran, R.A.J. Wharton, D.J. des Marais and C.P. McKay, J. Geophys. Res.-Planet. 103, 28481–28488 (1998).
- H.G.M. Edwards, “Raman Spectroscopic Analysis of Extremophilic Organisms under Martian Conditions”, in ROME: Response of Organisms to the Martian Environment, Ed by C.S. Cockell and G. Horneck. ESA Special Publication SP-1299 from the ESA ROME Topical Team 2002–2004, ESA Communications Publishing, Noordwijk, Netherlands, pp. 183–200 (2007).
- D.D. Wynn-Williams and H.G.M. Edwards, “Environmental UV-radiation: Biological strategies for protection and avoidance”, in Astrobiology: The Quest for the Origins of Life, Ed by G. Horneck and C. Baumstarck-Khan. Springer-Verlag, Berlin, pp. 245–260 (2002).
- D.D. Wynn-Williams, “Antarctica as a model for ancient Mars”, in The Search for Life on Mars, Ed by J.A. Hiscox. British Interplanetary Society, London, pp. 49–57 (1999).
- D.D. Wynn-Williams, “Cyanobacteria in deserts–life at the limit?”, in The Ecology of Cyanobacteria: Their Diversity in Time and Space, Ed by B.A. Whitton and M. Potts. Kluwer Academic Press, Dordrecht, The Netherlands, pp. 341–366 (1991).
- H.G.M. Edwards, Philos. T. Roy. Soc. A 368, 3059–3066 (2010).
- J.A. Brier, S.N. White and C.R. German, Philos. T. Roy. Soc. A 368, 3067–3086 (2010).
- E.A. Carter, M.D. Hargreaves, T.P. Kee, M.A. Pasek and H.G.M. Edwards, Philos. T. Roy. Soc. A 368, 3087–3098 (2010).
- H.G.M. Edwards, F. Sadooni, P. Vitek and J. Jehlicka, Philos. T. Roy. Soc. A 368, 3099–3108 (2010).
- J. Jehlicka, H.G.M. Edwards and A. Culka, Philos. T. Roy. Soc. A 368, 3109–3126 (2010).
- S.E. Jorge Villar and H.G.M. Edwards, Philos. T. Roy. Soc. A 368, 3127–3136 (2010).
- C.P. Marshall and A. Olcott Marshall, Philos. T. Roy. Soc. A 368, 3137–3146 (2010).
- P. Vitek, H.G.M. Edwards, J. Jehlicka, C. Ascaso, A. de los Rios, S. Valea, S.E. Jorge Villar, A.F. Davila and J. Wierzchos, Philos. T. Roy. Soc. A 368, 3205–3221 (2010).
- F. Rull, A. Delgado and J. Martinez-Frias, Philos. T. Roy. Soc. A 368, 3145–3152 (2010).
- F. Rull, M.J. Munoz-Espadas, R. Lunar and J. Martinez-Frias, Philos. T. Roy. Soc. A 368, 3153–3166 (2010).
- S.K. Sharma, A.K. Misra, S.M. Clegg, J.E. Barefield, R.C. Wiens and T. Acosta, Philos. T. Roy. Soc. A 368, 3167–3192 (2010).
- T. Varnali and H.G.M. Edwards, Philos. T. Roy. Soc. A 368, 3193–3204 (2010).
- H.G.M. Edwards, N.C. Russell and D.D. Wynn-Williams, J. Raman Spectrosc. 30, 685–690 (1997).
- H.G.M. Edwards, S.E. Jorge Villar, J.A. Parnell, C.S. Cockell and P. Lee, Analyst 130, 917–923 (2005).
- S.E. Jorge Villar and H.G.M. Edwards, Anal. Bioanal. Chem. 384, 1356–1365 (2006).
- H.G.M. Edwards, D.W. Farwell, M.M. Grady, D.D. Wynn-Williams and I.P. Wright, Planet. Space Sci. 47, 353–362 (1999).