Kit D. Jernshøj and Søren Hassing
University of Southern Denmark, Campusvej 55, DK-5230 Odense M, Denmark. E-mail: [email protected]
Introduction
A Raman spectrum reflects the vibrational motion of the nuclei within a molecule. When the wavenumber of the excitation laser is chosen to be close to that of an electronic absorption of a chromophore in a biomolecule, a resonance Raman spectrum is produced. Local molecular information is obtained since only those vibrations associated with the electronic transition in the chromophore are enhanced in the resonance Raman spectrum. The polarisation of the inelastically scattered light may be different from that of the incident laser light, the difference being determined by the nature of the nuclear vibrational motion. In resonance Raman scattering (RRS), the amount of structural and chemical information deduced can be increased by analysing the polarisation of the inelastically scattered light. The polarisation is defined by the depolarisation ratio (DPR), i.e. the ratio between the scattered intensity with perpendicular polarisation compared to the incoming light and that which is parallel polarised. Since the Raman process is described by a tensor (the polarisability), it is possible to gain additional molecular information from polarised measurements on randomly oriented systems such as solutions.1,2
The benefits of analysing the polarisation effects should be considered in contrast to using only the unpolarised resonance Raman spectra in a classification of molecular species, detecting an aggregation or conformational change. The decision is in the latter case (using only the unpolarised spectrum) based on either occasional small wavenumber shifts of the Raman modes and/or relative intensity changes between these, or the appearance of new bands in a spectrum. The decision based on the polarised spectrum is independent of these factors.
Another complementary technique used is dynamic light scattering (DLS), by which it is possible to obtain, for example, information regarding the macroscopic size (average diameter) of biological units. In DLS experiments, the intensity fluctuations of the elastically scattered light caused by the diffusion or Brownian motion of the scatterers are analysed. The intensity fluctuations can be converted into a size distribution by means of appropriate assumptions and mathematical tools as well as Mie theory.3
Aggregation of bio-molecules plays an important role in various bio-physical processes. The polarisation properties of RRS can reveal molecular changes such as aggregation and conformational changes; hence polarised Raman spectra can be used to study the degree of molecular aggregation in bio-molecules in their natural environment.
Revealing aggregated haemoglobin molecules in living erythrocytes
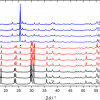
Haemoglobin (Hb) molecules with four oxygen binding sites assemble in erythrocytes in nearly all vertebrates. The aggregation of Hb molecules in living erythrocytes of human blood has been studied using polarised Raman scattering on spatially oriented erythrocytes combined with a principal component analysis (PCA).4 The conclusion reached was that a distinct ordering of haemoglobin took place within the erythrocytes. It is possible, however, to reach a similar conclusion regarding aggregation of intracellular Hb with a minimum of sample preparation and without orienting the erythrocytes. This can be achieved by taking advantage of the tensorial nature of the Raman process by measuring the polarised resonance Raman spectra of: (1) a dilute solution containing a number of randomly oriented erythrocytes (see Figure 1), and (2) a dilute reference solution containing a number of randomly oriented Hb molecules (not shown) and finally calculating the DPR values of the different modes (a2g modes) from these spectra.
, parallel polarised (red), 532 nm excitation] of a solution containing randomly oriented erythrocytes in phosphate buffered saline (PBS).")
In RRS, there is no upper limit of the DPR value; it may be either a constant or depend on the excitation wavenumber (polarisation dispersion). Figure 2 shows a model calculation giving the DPR (a2g mode) as a function of the wavenumber in the case of dimerisation of Hb. When an isolated haeme-group exhibits its ideal four-fold symmetry, the DPR of the a2g modes shows no dispersion and has the value infinity, but when encountering a symmetry lowering perturbation (for example, the incorporation into Hb), the DPR of these modes becomes dispersive and will decrease. In Reference 5, it is calculated that, for porcine Hb, the a2g mode n19 has the DPR value, DPRn19 = 7 ± 1.
 of an Hb dimer as a function of the wavenumber. The excitation wavenumber is 18.8 kcm–1.")
From model calculations accounting for intermolecular interactions between Hb molecules (monomers), it follows that the excited states of these individual molecules, upon aggregation, will split into a number of states with slightly different energies, see Figure 3.

Luckily, due to the nature of the Raman process, even small changes in the electronically excited states, which participate in the RRS process, will give rise to noticeable changes in the DPR. These changes are caused by alterations in the interference between the excited states participating in the resonance Raman process. The aggregation of Hb will, therefore, induce yet more changes in the polarisation dispersion of the dispersive a2g modes.
In practice, the DPR value needs to be even further decreased in order to unequivocally conclude that the Hb molecules may be aggregated. In the case of aggregated Hb in the erythrocytes, the DPR value is DPRn19 = 3 ± 1.5 Hence, it is possible to demonstrate an aggregation or perturbation by monitoring the polarisation properties in the form of changes in the DPR values.
A study of extracellular giant haemeoglobin in Arenicola marina based on polarised RRS and DLS
Extracellular haemoglobin (Hbl Hb) from Arenicola marina (lugworm) is considered to be a promising candidate as a blood substitute due to, for example, the high resistance towards oxidation of the haeme Fe ion. This is explained by the very complex, crowded and protected haeme environment. As opposed to the four oxygen-binding sites in vertebrate Hb, Hbl Hb has 144 haeme groups (or oxygen binding sites) and is freely dissolved within the vessels of the lugworm. The giant molecule has a molecular mass in the mega Dalton regime, i.e. approximately 4 × 106 Da.6,7,8 Biophysical and structural information about the giant protein can be disentangled by combining the unique polarisation properties of RRS with the size information obtained by DLS.
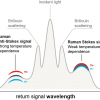
The DLS measurements on dilute solutions of Hbl Hb, which provides an average hydrodynamic diameter, dh, of Hbl Hb, were performed at several pH values.5 At pH = 5 to 7, the Hbl Hb molecule exists in its native form, in which case the average diameter, dh, is equal to 24.2 nm. At pH 9, a dissociation process has taken place, resulting in dh = 9 nm, see Figure 4. At pH = 10, however, only large aggregates, which consist of fragmented Hbl Hb, with dh larger than 1000 nm, could be detected.

A comparison of the DLS results for pH = 10, with the behaviour of the polarised RRS measurements (the DPR values of three a2g modes), reveals that no further distortion of the haeme group took place upon aggregation. Hence, the coupling between the fragments or aggregation did not involve any direct interaction between the haeme groups.
 and perpendicularly polarised (blue)] of native Hbl Hb from Arenicola marina (pH = 5), obtained with 532 nm excitation. Reproduced with permission from J. Chem. Phys. 139, 065104 (2013). Copyright 2013 American Institute of Physics.")
In addition, an analysis of the DPR of three a2g modes of the haeme in native Hbl Hb and porcine Hb shows that the distortion of the haeme group away from its ideal four-fold symmetry is smaller for haeme groups bound in the lugworm Hbl Hb (i.e. DPRn19 = 15 ± 2) than for haeme groups bound in porcine Hb (DPRn19 = 7 ± 1). To summarise:
Molecule | DPRn19 |
Haeme group | ∞ |
Porcine Hb | 7 ± 1 |
Porcine Hb aggregated in erythrocytes | 3 ± 1 |
Hbl Hb | 15 ± 2 |
The visible absorption spectra of porcine Hb, erythrocytes and Hbl Hb (pH = 5 and 10) are alike, except for minor differences not relevant here, which indicates that the molecular configuration of the haeme group corresponding to the excited state is not strongly perturbed. By analysing the polarisation properties, it is possible to gain additional molecular information sometimes barely accesible using the unpolarised spectra.
Outlook
In a forthcoming publication, a resonance Raman-based polarisation study of the thermal stability of a dye from dye-sensitised solar cells is reported. One of the key parameters governing the long-term thermal stability of a dye-sensitised solar cell is, of course, the dye stability, i.e. whether or not this is degraded by thermal stress.
Acknowledgement
K.D. Jernshøj greatly acknowledges support from Energi Fyn.
References
- D.A. Long, The Raman Effect: A Unified Treatment of the Theory of Raman Scattering by Molecules. John Wiley & Sons Ltd, Chichester, UK (2002).
- S. Hassing and O. Sonnich Mortensen, Advances in Infrared and Raman Spectroscopy, Vol. 6: Polarization and Interference Phenomena in Resonance Raman Scattering. Heyden and Sons, London, UK (1980).
- R. Pecora (Ed), Dynamic Light Scattering, Applications of Photon Correlation Spectroscopy. Plenum Press, New York, USA (1985).
- B.R. Wood, L. Hammer and D. McNaughton, “Resonance Raman spectroscopy provides evidence of heme ordering within the functional erythrocyte”, Vib. Spectrosc. 38(1–2), 71 (2005). doi: 10.1016/j.vibspec.2005.02.016
- K.D. Jernshøj, S. Hassing and L.F. Olsen, “A combination of dynamic light scattering and polarized resonance Raman scattering applied in the study of Arenicola marina extracellular hemoglobin”, J. Chem. Phys. 139(6), 065104 (2013). doi: 10.1063/1.4813920
- L.M. Moreira, A.L. Poli, J.P. Lyon, F.V. Santos, P.C.G. Moraes, J.P.F. Mendonça, V.C. Barbosa and H. Imasato, “Giant extracellular hemoglobin of Glossoscolex paulistus: excellent prototype of biosensor and blood substitute”, Chapter 18, in Biosensors for Health, Environment and Biosecurity. InTech Open Access Publisher (2011).
- M. Tabak, F.A.O. Carvalho, J.W.P. Carvalho, J.F.R. Bachega and P.S. Santiago, “Recent characterizations on the giant extracellular hemoglobin of Glossoscolex paulistus and some other giant hemoglobins from different worms, Chapter 15, in Stoichiometry and Research—The Importance of Quantity in Biomedicine, InTech Open Access Publisher (2012).
- J. Elmer and A.F. Palmer, “Biophysical properties of Lumbricus terrestris erythrocruorin and Its potential use as a red blood cell substitute, Open Access J. Funct. Biomater. 3(1), 49 (2012). doi: 10.3390/jfb3010049