Martin Byrdin and Dominique Bourgeois
Institut de Biologie Structurale, Univ. Grenoble Alpes, CEA, CNRS, F-38044 Grenoble, France
Introduction
Microspectrometers are (sometimes bulky) spectrometers specifically designed for the study of microscopic samples. A variety of instruments have been developed that combine spot sizes down to sub-microns with spectral ranges from ultraviolet (UV) to infrared (IR) for various optical spectroscopy modalities such as absorbance, transmittance, fluorescence or vibrational spectroscopies. If, on top of being tiny, the samples are delicate objects such as protein microcrystals, other challenges than just tight and achromatic focusing arise: sample optical anisotropy, large refractive index and demanding environmental requirements make most commercially available devices unsuited. Therefore, at many places where microspectroscopy is required (for example at synchrotron sources to investigate protein crystals), home-made microspectrometers have been developed in recent years that strive to meet the above-mentioned challenges.1
At the European Synchrotron Radiation Facility in Grenoble, France (ESRF), a dedicated system based on three mutually aligned mirror objectives has been installed (“Cryobench”)2,3 that combines the absorption (A), fluorescence (F) and Raman modalities with the possibility of rotational adjustment of the sample (via a goniometric support), and the option to cool the sample down to 100 K due to a stream of gaseous nitrogen. Our experience with the powerful possibilities of this system, and the realisation of its limitations prompted us to design a next generation microspectrometer dubbed “CAL(AI)2DOSCOPE” which is presented in this article. The biological samples we study (fluorescent proteins used as genetically encoded markers for advanced fluorescence microscopy) exhibit an intricate behaviour highly sensitive to their micro-environment and notably to their illumination history. As a consequence, their absorption and fluorescence (giving access to their photophysical properties) cannot be studied independently, but rather need to be followed (quasi-) simultaneously, requiring coordination in space and time. Existing solutions such as crossed beams (e.g. ESRF Cryobench)4 or optics rearrangement (e.g. SwissLightSource)5 are not fully compatible with these requirements. Therefore, we developed the alternative solution of a common optical path for both A and F, coupled to rapid switching of light sources and detectors by upstream mechanical shutters.
Design features of the CAL(AI)2DOSCOPE
i) The CAL(AI)2DOSCOPE was designed and fabricated by the opto-mechanics company Optic Peter (www.optiquepeter.com, France). The key feature of the instrument is an optical layout in which light beams for absorption, fluorescence and actinic illumination are arranged colinearly and pass through a common objective. This layout is inspired by the design of classical epi-fluorescence microscopes (see Figure 1 for a schematic view). It provides a solution to the complex alignment problems encountered with non-colinear beams, for which insertion of each new sample of finite thickness and specific refractive index compromises beam overlap (displacement, inclination) and hence requires reajustment. The use of a common objective for diverse excitation/observation tasks simplifies the issue of mutual excitation/observation alignment to that of proper tuning of the upstream semi-transparent beam coupling optics. Alignment of F and A thus becomes essentially sample-independent and can be accomplished once and for all before any measurement series. Measurement of identical sample portions for A and F is thus optimised by design and light-induced colour changes or residual minor mechanical instabilities at the sample level no longer impact overlap.

Figure 1. Opto-mechanical setup of the CAL(AI)2DOSCOPE. The four light-blue knobs (x, y, z, θ) serve to position the sample with respect to the fixed top objective, the three dark-blue knobs serve to position the lower objective with respect to the sample (and hence the top objective).
ii) Although looking like a microscope from the outside (Figure 2), our device has features that are optimised for a spectrometer, notably to minimise chromatic aberrations. Thus, objectives are mirror objectives (Cassegrain design), the fibre collimators are off-axis parabolic mirrors and the beam splitters are optical sieves (polka dot design), ensuring a flat spectral response. In total, the absorption light path contains no less than seven mirrors: one per fibre coupling (×2), two per objective (×2), one per beamsplitter (×1), and consequently the overall optical throughput does not exceed a few percent.

Figure 2. View of the instrument. A: Overall view: the microscope stand is on the left, the laser assembly on the right, the timing electronics on the top and the cryosystem is under the optical table. Of the two computer screens, the top one shows the camera image of the sample (for alignment) and the one on the right shows a kinetic trace from the fluorescence spectrometer. B: Sketch of the CAL(AI)2DOSCOPE. C: Zoom into sample space: the space between the symmetric Cassegrain objectives is large enough to house the goniometric sample holder and the cryonozzle coming from the left. (The piece is 2€ with diameter 25 mm.)
iii) The highly rigid microscope stand ensures that the pre-aligned positions of the objectives and samples are kept stable over time. The stand does not incorporate light sources or detectors, except a camera for the observation of the alignment process and the documentation of its result. Instead, it houses four entrance/exit gates that can accommodate at will fibre-coupled devices such as two CCD spectrometers or any desired light source. Beside tungsten and halogen lamps, several diode-pumped solid-state (DPSS) lasers at selected wavelengths between 405 nm and 561 nm are aligned to a common path and can provide stable measuring/actinic light. Between each of the microscope’s fibre ports and its common main optical axis, there is space for the insertion of 1-inch circular plates such as filters or polarisers.
iv) It is clear that for different excitation/detection wavelengths, different dichroic beam splitters are required and for different partition of light between the absorption and fluorescence paths, different reflectivities of the beam splitters are desired. In order to achieve this flexibility and to maintain the advantages of the pre-aligned layout, we introduced, again inspired by microscope design, four-position stages at the level of each of the beam splitters, thus allowing for one-click repositioning between pre-aligned elements via a slide bar (see Figure 2).
v) To avoid mutual perturbation between the A and F modalities, we use alternate detection. This is to say that at any time there is either laser illumination and fluorescence detection ON (and absorbance lamp and spectrometer OFF) or vice versa. To this end, mechanical shutters allow for temporal synchronisation between detectors and sources. The shutters are synchronised via a Labview interface on the measuring computer that also receives the data from the spectrometers. Temporal resolution is limited in theory by the opening time of the shutters (about 1 ms), in practice more often by the integration time needed to obtain reasonable signal-to-noise ratios on the spectrometer chips (more like 10 ms). As a consequence, photo-physical processes that evolve at rates faster than ~100 Hz are currently beyond detectability. The quasi-synchronous detection opens the possibility to study the time course of photo-reactions in several types of immobilised samples, e.g. crystalline, liquid or embedded in polymer matrices, opening the way to a broad range of applications.
vi) In the actual layout, a significant part of the measuring light is lost, due to the presence of the beam splitters in the common optical path. An obvious improvement both in time resolution (switching speed) and signal amplitude (light intensity) could be obtained by using for A/F beam separation a rotating mirror or an acousto-optical device instead of a beam splitter, so that the totality of the available light could reach the active detector (both solutions do in principle exist, but would require further adaptation of the mechanical design). An important desirable development concerns automated sample positioning, possibly with the option of multi-well plate scanning, opening the door to high-throughput screening of multiple samples, e.g. variants of fluorescent proteins produced by directed evolution approaches.
Application
We look at the case of phototransformable fluorescent proteins: why do we need to study these proteins under different phases (crystalline versus solution), with A/F at the same spot at the same time, and, moreover, under actinic illumination?
>p>The proteins that are studied in our group are so-called “phototransformable fluorescent proteins” (PTFPs): their A/F properties are sensitive to light and change with illumination, a phenomenon known in absorbance as photochromism. For many practical applications in bioimaging or biotechnology, light-induced changes in fluorescence (excitation/emission wavelength, quantum yield) are crucial. For example, fluorescence that can be optically induced, stopped or even toggled between ON and OFF states, provides a handle to distinguish individual chromophores (and important intracellular biological molecules labelled by them) and opens the door for advances in particle tracking, interaction studies or super-resolution fluorescence microscopy… Many of these exciting techniques are made possible by the switchability of protein fluorescence by actinic light, but the performances of these methods depend crucially on a precise understanding of the photo-physical processes exhibited by the fluorescent markers. This presents a considerable challenge and is the reason why our CAL(AI)2DOSCOPE has to be able to follow as many parameters as possible simultaneously on one and the same protein sample spot: the one that is hit by the actinic beam causing the photophysics to happen. Intrinsic timescales of photophysical processes within PTFPs range from (sub)nanoseconds to hours. However, time-scales relevant to advanced microscopy are typically those accessible by the CAL(AI)2DOSCOPE. Photoswitching or photobleaching are examples of photophysical processes, the rates of which can be conveniently investigated in this way.6,7

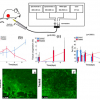
Figure 3. “Photofatigue” of a variant of the photoswitchable fluorescent protein mEos2 under OFF/ON cycles of actinic 405 nm light (violet bars) and permanent 488 nm fluorescence excitation (cyan bar). Panel A shows the temporal evolution of absorbance (around 490 nm, orange) and fluorescence (around 512 nm, olive) of a microdrop of an mEos2 mutant immobilised in a matrix of polyvinyl alcohol (PVA). The amplitudes are normalised for presentation on a common scale. Panel B shows the crystallographic structure of mEos2, the parent fluorescent protein with the green “barrel” structure housing the chromophore in red.
Figure 3 shows an example study of a variant of the phototransformable fluorescent protein mEos2. Absorption at 490 nm and fluorescence at 512 nm are detected simultaneously under constant 488 nm actinic illumination superimposed to alternate light of 405 nm causing fluorescence OFF and ON switching. With advancing switching cycles, the amplitude and the ON/OFF contrast diminish, a phenomenon known as “fatigue”. It can be seen that during OFF-switching, the fluorescence decays in a biphasic manner that differs from that of absorption, suggesting that the OFF-state, though not fluorescent, does still absorb at 490 nm. Another intriguing observation is that, towards the end of the ON-switching periods, fluorescence starts to diminish while absorption continues to rise. This subtle photophysical behaviour (hinting at a photobleaching mechanism induced by 405 nm light that now needs to be investigated in detail) would be difficult to detect with a standard apparatus and highlights the potential of the CAL(AI)2DOSCOPE.
Conclusion
The CAL(AI)2DOSCOPE incorporates in a single instrument multiple features that can only be found separately elsewhere:
- microscopic samples mountable on a motorised goniometric sample holder;
- camera-supported alignment with up to 15× optical magnification (numerically unlimited);
- quasi-simultaneous absorption and fluorescence spectroscopy on identical sample sections of down to 10< µm diameter;
- access to light-induced reactions (photo-physics, photo-chemistry) via programmable illumination of the same spot with various actinic sources; and
- controlled environment via continuous flow of gas of choice (air, argon, N2) at temperature of choice for the latter (100–300 K).
Overall, the modular design of the CAL(AI)2DOSCOPE allows for a maximum of versatility, adaptability and flexibility.
Although originally developed to investigate fluorescent protein photophysics, the instrument, thanks to its unique combination of features, should be useful in a much wider range of biological-chemical-physical-technological applications.
Acknowledgements
We thank the Optique Peter company for crucial help in the design and for the fabrication of the CAL(AI)2DOSCOPE. Elke de Zitter (KU Leuven) is acknowledged for providing the fluorescent protein sample discussed in the paper. We are grateful to Virgile Adam and Romain Berardozzi for inspiring discussions. D.B. acknowledges financial support by the ANR (ANR-2011-BSV5-012-01 NOBLEACH).
References
- A.R. Pearson, A. Mozzarelli and G.L. Rossi, “Microspectrophotometry for structural enzymology”, Curr. Opin. Struct. Biol.14, 656–662 (2004). doi: http://dx.doi.org/10.1016/j.sbi.2004.10.007
- D. Bourgeois, X. Vernede, V. Adam, E. Fioravanti and T. Ursby, “A microspectrophotometer for UV-visible absorption and fluorescence studies of protein crystals”, J. Appl. Crystallogr. 35, 319–326 (2002). doi: http://dx.doi.org/10.1107/s0021889802003837
- D. von Stetten, T. Giraud, P. Carpentier, F. Sever, M. Terrien, F. Dobias, D.H. Juers, D. Flot, C. Mueller-Dieckmann, G.A. Leonard, D. de Sanctis and A. Royant, “In crystallo optical spectroscopy (ICOS) as a complementary tool on the macromolecular crystallography beamlines of the ESRF”, Acta Crystallogr. D 71, 15–26 (2015). doi: http://dx.doi.org/10.1107/S139900471401517X
- T. Barros, A. Royant, J. Standfuss, A. Dreuw and W. Kühlbrandt, “Crystal structure of plant light-harvesting complex shows the active, energy-transmitting state”, EMBO J. 28, 298–306 (2009). doi: http://dx.doi.org/10.1038/emboj.2008.276
- R.L. Owen, A.R. Pearson, A. Meents, P. Boehler, V. Thominet and C. Schulze-Briese, “A new on-axis multimode spectrometer for the macromolecular crystallography beamlines of the Swiss light source”, J. Synchrotron Radiat. 16, 173–182 (2009). doi: http://dx.doi.org/10.1107/s0909049508040120
- C. Duan, V. Adam, M. Byrdin, J. Ridard, S. Kieffer-Jaquinod, C. Morlot, D. Arcizet, I. Demachy and D. Bourgeois, “Structural evidence for a two-regime photobleaching mechanism in a reversibly switchable fluorescent protein”, J. Am. Chem. Soc. 135, 15,841–15,850 (2013). doi: http://dx.doi.org/10.1021/ja406860e
- R. Berardozzi, V. Adam, A. Martins and D. Bourgeois, “Arginine 66 controls dark-state formation in green-to-red photoconvertible fluorescent proteins”, J. Am. Chem. Soc. 138, 558–565 (2016). doi: http://dx.doi.org/10.1021/jacs.5b09923